
Are There Any Cladistic Analyses of Malawi Cichlids?
![]() he first explicit cladistic analysis of any group of Malawi cichlids was that of Nimbochromis ("the Cyrtocara livingstonii species-group") in Oliver (1984). Although Eccles and
Trewavas (1989: 282) briefly discussed his analysis, they misunderstood and misrepresented the evidence for the group's monophyly. Oliver's cladogram, slightly modified, is given here with added thumbnail photos to facilitate comparison of the
larger features of the color pattern. The character evidence is summarized below the diagram.
he first explicit cladistic analysis of any group of Malawi cichlids was that of Nimbochromis ("the Cyrtocara livingstonii species-group") in Oliver (1984). Although Eccles and
Trewavas (1989: 282) briefly discussed his analysis, they misunderstood and misrepresented the evidence for the group's monophyly. Oliver's cladogram, slightly modified, is given here with added thumbnail photos to facilitate comparison of the
larger features of the color pattern. The character evidence is summarized below the diagram.
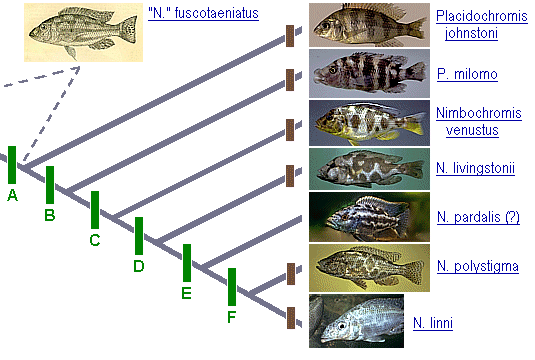
Shared derived characters (green bars)
Node A (including all species shown except N. fuscotaeniatus): 1. Color pattern of body consisting primarily of six broad, vertical bars, four of them situated partly or entirely below the dorsal-fin base as seen in
Placidochromis johnstoni [not "six well-marked spots along the dorsum at the base of the dorsal fin" as Eccles & Trewavas (1989: 282) claimed that I had stated]. This condition represents a reduction from the apparent primitive number of bars in
haplochromines (approx. 7-10), as well as an increase in the relative width and intensity of each bar. In exceptional individuals, the number of bars on the body is further reduced to five. Character 1 is further modified within subsets of the
species-group as described below (characters 5 and 7-10).
Within node A, the following synapomorphies suggest that all species except P. johnstoni form a monophyletic subunit, node B:
Node B: 2. Lower pharyngeal bone widened compared to that of P. johnstoni. 3. Definitive adult jaw teeth unicuspid (attained in fishes <120 mm SL in most species). 4. Maximum adult size increased from approx. 140 mm SL to at least 165 mm
SL. 5. Vertical bars on body somewhat broken or dorsally incomplete in many individuals of P. milomo and invariably broken into spots in the remaining species (see character 7).
Within node B, all species except P. milomo constitute a monophyletic group (node C) characterized by the following synapomorphies:
Node C (Nimbochromis sensu Eccles & Trewavas): 6. Pharyngeal teeth all compressed, cuspidate, the posteromedian teeth never having rounded, submolariform to molariform crowns; associated with shift from an insectivorous to a piscivorous
diet. 7. Some bars on flanks invariably broken into oblique series of 2 or 3 blotches.
Within node C, one synapomorphy suggests that all species except N. venustus form a monophyletic subgroup:
Node D: 8. Pectoral-fin rays with small dark spots. (Although I have not examined the holotypes of the nominal species N. pardalis and N. maculimanus, both their original descriptions and the accounts in Eccles & Trewavas mention
that they have small spots on the pectoral fins. Spotted pectorals are, I think, unique to node D among cichlids and highly unusual among teleosts.)
Within node D, N. pardalis, N. polystigma, and N. linni are a monophyletic subgroup characterized by the following synapomorphy which is absent in N. livingstonii:
Node E: 9. Body and head with small dark "freckles" superimposed on the pattern of blotches, the freckles distributed one to each scale over most of the body. Freckles may be indistinct in sexually active males.
Within node E, at least N. polystigma and N. linni share a further synapomorphy whose occurrence in N. pardalis is unknown at this time:
Node F: 10. Pelvic fins with several distinct, brown oval spots (masked in breeding males). Spotted pelvic fins are unique to this node within the Cichlidae, so far as I know.
One implication of this analysis, if correct, is that Placidochromis including P. johnstoni and P. milomo, as constituted by Eccles and Trewavas (1989), is not a monophyletic group.
Unique derived characters (autapomorphies) of each species (brown bars)
The following characters either appear to be unique to a particular member of this group, or are most parsimoniously interpreted as independently derived within a member of the group and in one or more species outside the group. Most of the osteological
characters are illustrated in Oliver (1984).
P. johnstoni: (a) Premaxillary symphysis pointed. Since the sister group of this entire group is still unknown, this character may prove to be more widely distributed.
P. milomo: (b) Lips greatly hypertrophied and medially lobate; (c) maxilla strongly curved; (d) dentary short and deep; (e) gill rakers on lower arm of outer arch increased to 13-15 (from a range of 10-12[-13]). Hypertrophied lips are known
in several other haplochromines. Within Lake Malawi, the most extreme hypertrophy outside that in P. milomo is found in Abactochromis labrosus of the mbuna flock. Because of the different color pattern in Eclectochromis species,
which have plesiomorphic (more numerous and narrower) vertical bars, I interpret the less hypertrophied lips in that genus as a convergent character.
N. venustus: (f) Caudal fin lacking the dark-edged, orange maculae that are primitively present in this group; (g) ventral half of caudal fin yellow (this coloration masked in adult males); (h) breeding males with sulphur-yellow median blaze
on nape; (i) dentary, in lateral view, tapering anteriorly to shallow symphysis; (j) anterior dentary teeth procumbent.
N. livingstonii: (k) Ground color of body chalky white in life; (l) distinctive feeding behavior involving behavioral mimicry of a dead fish by dropping to substrate, lying motionless on its side to lure small fishes as prey (Fryer & Iles,
1972; McKaye, 1981).
[N. pardalis: No autapomorphies known]
N. polystigma: (m) Maxilla nearly straight.
N. linni: (n) Snout elongate; (o) mouth subterminal; (p) dentary dorsoventrally flattened; (q) ventral profile of retroarticular and anguloarticular forming an obtuse angle (instead of a straight line) with ventral profile of dentary; (r)
mesopterygoid much longer than deep; (s) maxillary process of palatine elongate; (t) premaxilla with ascending process longer than alveolar process, and angle between the two processes reduced; (u) neurocranium elongate (reflected in reduced otic depth
and reduced angle of preorbital profile of skull); (v) distinctive feeding behavior, involving ambush of small fishes from behind a rock with snout resting on top of rock.
In both flocks, moreover, most species have quite uniform skeletal morphology — as might be expected in groups that speciated rapidly in a relatively short time. For example, within the larger, or "Hap" flock, Oliver found no discrete interspecific
variation in the number or configuration of the caudal skeleton elements (although considerable intraspecific variation occurs); no easily used variation in the gill arch skeleton (although numbers of gill rakers, of course, vary inter- and
intraspecifically); the same bones in the neurocranium of every species examined (although proportions of bones and bone complexes certainly do vary, as exploited in the mbuna flock by Reinthal); little variation in the pelvic and pectoral skeleton; and
so on.
What remains are harder-to-manage continuous variables that often overlap interspecifically and change ontogenetically, such as bone proportions and body proportions, as well as color patterns, ranges of counts, behavioral traits, enzymes, and genes.
DNA sequences may provide the ultimate characters for cladistic analysis — but they will need to be interpreted using the same methods of character polarity determination with out-groups as any other cladistic data, grouping taxa using shared, derived
characters (synapomorphies). "Common" within the group cannot be equated with "primitive." [If it could, viviparity, not oviparity, would be the primitive reproductive mode of mammals!]
This is not to say that cladistic analysis and classification of these cichlids is impossible or pointless. It needs to be done; it can be done, as shown by the above small example; it will eventually be accomplished for all species. It will not come
easily. Yet, only a classification based on tested and corroborated phylogenetic hypotheses can hope to approach stability. Only a cladistic classification will offer scientists and all interested persons the benefits of summarizing all our knowledge
about these fishes, of predicting the distribution of newly discovered characters, and of reflecting our best estimate of the single actual (even though never known with certainty) phylogenetic history of each group.
![]() ladistic analysis of Malawi haplochromines is exceptionally difficult. The sheer number of taxa is daunting, making truly comprehensive character surveys nearly impossible and the assembly of study material of even representative species a
challenge.
ladistic analysis of Malawi haplochromines is exceptionally difficult. The sheer number of taxa is daunting, making truly comprehensive character surveys nearly impossible and the assembly of study material of even representative species a
challenge.





Last Update: 29 October 2010
Web Author: M. K. Oliver, Ph.D.
Copyright © 1997-2021 by M. K. Oliver, Ph.D. - ALL RIGHTS RESERVED